Hvad præcise størrelsesmålinger af proteiner kan fortælle os om deres foldning, binding og hvordan de samles. Fra Einsteins diffusionslov og Taylor-dispersion til moderne kapillærmetoder er proteinstørrelse blevet et overraskende stærkt mål for binding, aggregering og selvsamling.
Artiklen har været bragt i Dansk Kemi nr. 3, 2026 og kan læses uden illustrationer, strukturer eller ligninger herunder
(læs originalartiklen her)
Af Alexander Kai Büll, Institut for Bioteknologi og Biomedicin, DTU
Proteiner og deres interaktioner er vigtige på områder, som umiddelbart virker langt fra hinanden: i lægemiddeludvikling, hvor man vil vide, om et terapeutisk antistof faktisk når og binder til virkningspunktet, og i fødevarevidenskab, hvor proteiners måde at samle sig på styrer tekstur, stabilitet og funktion. Derfor er det værd at se nærmere på en tilsyneladende enkel egenskab: størrelse.
Ved første øjekast lyder det næsten naivt. Proteiner er kemisk komplekse og strukturelt dynamiske molekyler, så hvorfor skulle noget så “groft” som den samlede størrelse fortælle os noget nyttigt? Svaret er, at præcise størrelsesmålinger i opløsning ofte er overraskende informative. Når et protein folder sig, binder en partner, danner oligomerer, kondenserer eller aggregerer, ændrer det sin størrelse – og dermed også sin diffusion. Hvis vi kan måle den diffusion præcist nok, bliver proteiners størrelse et følsomt vindue ind til molekylær adfærd.
En kort historie om, hvorfor størrelse betyder noget
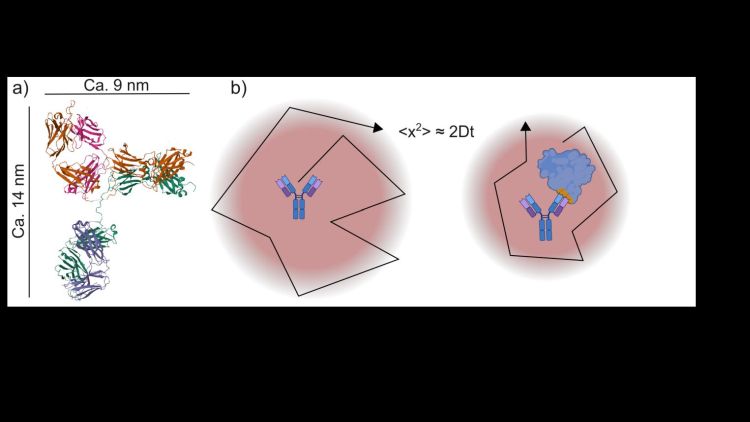
I begyndelsen af det 20. århundrede vidste forskere endnu ikke, om proteiner var selvstændige molekyler med veldefinerede masser og størrelser, eller blot et uklart kolloidalt materiale. Theodor Svedbergs ultracentrifuge, udviklet i Uppsala, gjorde det muligt at bestemme molekylvægte og var med til at fastslå, at proteiner er ægte og veldefinerede makromolekyler (det vil sige molekyler, der består af tusindvis af atomer) med karakteristiske størrelser (figur 1 a).
Omtrent samtidig gav Einsteins forklaring på Brownske bevægelser diffusion et solidt teoretisk grundlag. Gennem Stokes-Einstein relationen kan diffusionskoefficienten D for en kugleformet partikel eller et molekyle kobles direkte til radius R (figur 1 b), når opløsningens viskositet η er kendt (T er temperaturen, og kB er Boltzmanns konstant):
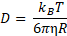
Sagt på en anden måde: diffusion – den måde, molekyler bevæger sig på som følge af termiske fluktuationer – blev en vej til at måle, hvor stort et molekyle er [1].
Det næste vigtige skridt mod den teknologi, vi taler om her, kom fra fluidmekanikken. I 1950’erne viste Geoffrey Taylor [2] og Rutherford Aris [3], at når et lille volumen af et opløst stof bevæger sig gennem et smalt rør under laminær strømning, så spredes det, fordi væsken bevæger sig hurtigere i midten end tæt ved væggen (figur 2 a). Diffusion virker på tværs af rørets radius og udjævner delvist disse hastighedsforskelle, så der opstår et karakteristisk spredningsmønster, der ligner en Gaussisk funktion med variansen σ² (figur 2 b+c). Tilsammen skabte disse studier grundlaget for at udlede diffusionskoefficienter ud fra bredden af denne Gauss-profil (D ∝ 1/σ²) og dermed molekylers størrelse ud fra peak-bredning under flow i kapillærer – det fysiske grundlag for Taylor-dispersionsanalyse, TDA.
Fra Taylor-dispersion til flowinduceret dispersionsanalyse
Den klassiske idé har fået en moderne biologisk anvendelse i flowinduceret dispersionsanalyse, som normalt forkortes FIDA. Hervé Cottets gruppe i Montpellier havde vist, at TDA kunne give præcis information om størrelsen af makromolekyler [4].
Henrik Jensen og Jesper Østergaard fra Københavns Universitet (KU) demonstrerede derefter styrken ved proteinanalyse med fluorescens som detektionsmetode og gav denne anvendelse navnet FIDA [5]. Det førte siden til kommercialisering af teknologien gennem en spinoutvirksomhed fra KU, FIDA Biosystems ApS, Fidabio. Konceptuelt lignede det oprindelige instrument en opstilling til kapillærelektroforese [4], men uden det elektriske felt. I stedet driver en kontrolleret trykforskel prøven gennem kapillaret. Detektionen er fluorescensbaseret, hvilket gør metoden meget følsom – koncentrationer i nanomolarområdet kan måles uden problemer.
At FIDA gør det muligt at måle proteininteraktioner i fri opløsning, står i kontrast til en af de vigtigste alternative metoder, nemlig overfladeplasmonresonans (surface plasmon resonance, SPR). Den metode kræver, at proteinerne bindes til en overflade, hvilket giver sine egne udfordringer, men til gengæld gør det let at måle, hvor hurtigt binding opstår og brydes. Det er for nylig også vist, at FIDA kan måle denne type kinetik direkte i opløsning [6].
FIDA-platformen er attraktiv, fordi den både er enkel og fleksibel. Arbejdsgangen er automatiseret, og prøveforbruget er meget lavt – 1 milliliter ville være nok til mindst ti tusind målinger. Brugeren kan desuden justere vigtige parametre som drivtryk, kapillærens dimensioner, overfladebelægning og detektionsbølgelængde. Fidabio er den førende, men ikke den eneste kommercielle leverandør af diffusionsbaseret størrelsesmåling. Beslægtede metoder tilbydes også af Fluidic Sciences i Storbritannien og Nanoscale Metrix i Frankrig – et tegn på metodens potentiale og voksende betydning i akademisk og industriel biomolekylær forskning.
Aflæsning af peaket
En af grundene til, at metoden er så nyttig, er, at selve signalets form indeholder information. Små partikler, der diffunderer hurtigt, giver et næsten symmetrisk, Gauss-formet peak, når de når detektoren (figur 2 b+c). Blandinger af forskellige diffunderende komponenter kan matematisk skilles ad med forskellige former for dataanalyse (figur 3 b) [7,8]. Bredere eller skæve peaks kan pege på større eller mere klæbrige komponenter (figur 3 b), mens meget store partikler, herunder aggregater på mikrometerskala, viser sig som enkelte skarpe signaler snarere end som glatte fordelinger (figur 3 c) [9,10].
Et enkelt kapillærforsøg kan derfor ikke kun fortælle os den gennemsnitlige størrelse, men også om en prøve er homogen, heterogen eller domineret af store aggregater.
Det er vigtigt, fordi mange biologisk vigtige hændelser direkte giver ændringer i størrelse. Binding mellem et antistof og et antigen [11,12] øger for eksempel størrelsen af den målte partikel (figur 3 a). Delvis udfoldning kan blotlægge klæbrige områder og få molekylerne til at binde sig til hinanden [13]. Størrelse er derfor ikke bare et groft mål; det er et samlet signal, som fortæller os, hvad proteiner gør i opløsning.
Interaktioner i rene og komplekse prøver
De første anvendelser af FIDA fokuserede på at kvantificere ikke-kovalente interaktioner i meget små prøver, og metoden er stadig stærk til enkle affinitetsmålinger i rene prøver (figur 3 a). Men feltet er gået langt videre end enkle målinger af binding mellem to partnere. FIDA-baserede arbejdsgange er blevet brugt til at kvantificere proteiner direkte i plasma og til at screene proteiner designet med kunstig intelligens direkte i urensede celleekstrakter (figur 3 d) [14]. Selv binding af små molekyler til proteiner, som ikke giver en målbar ændring i størrelse, kan kvantificeres gennem effekten på fluorescensspektret [15]. Parallelt har kapillær-flowmetoder fra vores gruppe på DTU udvidet den samme fysiske ramme til biomolekylære kondensater [9,16], amyloidsystemer [8] og fødevarerelevante caseinmiceller (figur 3 c) [9].
Disse eksempler viser en vigtig pointe: metodens egentlige styrke er ikke kun, at den kan fortælle os, om to molekyler interagerer, men at den kan følge, hvordan proteiner samler sig i heterogene og funktionelle tilstande.
For både bioteknologi og fødevarevidenskab er det et stærkt perspektiv. Proteiner optræder sjældent som isolerede monomerer. De binder, skifter partnere, danner klynger, faseadskilles og bygger større strukturer. Præcise størrelsesmålinger erstatter ikke højopløselige strukturmetoder, men de supplerer dem ved at give direkte information i opløsning, under forhold der ligner de naturlige, og med meget små prøvemængder. Nogle gange viser det sig, at et enkelt spørgsmål til et protein – “hvor stort er du lige nu?” – kan afsløre overraskende meget om, hvordan det folder sig, binder og samles.
E-mail:
Alexander Kai Büll: alebu@dtu.dk
Referencer
1. Otzen, D.E., Buell, A.K. and Jensen, H. (2021) Microfluidics and the quantification of biomolecular interactions. Curr. Opin. Struct. Biol. 70, 8-15. https://doi.org/10.1016/j.sbi.2021.02.006.
2. Taylor, G.I. (1953) Dispersion of soluble matter in solvent flowing slowly through a tube. Proc. R. Soc. Lond. A Math. Phys. Sci. 219, 186-203. https://doi.org/10.1098/rspa.1953.0139.
3. Aris, R. (1956) On the dispersion of a solute in a fluid flowing through a tube. Proceedings of the Royal Society of London. Series A 235, 67-77. https://doi.org/10.1098/rspa.1956.0065.
4. Cottet, H., Biron, J.P. and Martin, M. (2007) Taylor dispersion analysis of mixtures. Anal. Chem. 79, 9066-9073 https://doi.org/10.1021/ac071018w.
5. Jensen, H. and Østergaard, J. (2010) Flow induced dispersion analysis quantifies noncovalent interactions in nanoliter samples. J. Am. Chem. Soc. 132, 4070-4071. https://doi.org/10.1021/ja100484d.
6. Willmer, P., Stender, E.G.P., Sahni Ray, K., Hundahl, A.C., Marie, R. and Jensen, H. (2025) In-Solution Characterization of Biomolecular Interaction Kinetics under Native Conditions. Anal. Chem. 97, 19498-19504. https://doi.org/10.1021/acs.analchem.5c02164.
7. Cipelletti, L., Biron, J.P., Martin, M. and Cottet, H. (2015) Measuring Arbitrary Diffusion Coefficient Distributions of Nano-Objects by Taylor Dispersion Analysis. Anal. Chem. 87, 8489-8496. https://doi.org/10.1021/acs.analchem.5b02053.
8. Farzadfard, A., Kunka, A., Mason, T.O., Larsen, J.A., Norrild, R.K., Dominguez, E.T., et al. (2024) Thermodynamic characterization of amyloid polymorphism by microfluidic transient incomplete separation. Chem. Sci. 15, 2528-2544. https://doi.org/10.1039/d3sc05371g.
9. Stender, E.G.P., Ray, S., Norrild, R.K., Larsen, J.A., Petersen, D., Farzadfard, A., et al. (2021) Capillary flow experiments for thermodynamic and kinetic characterization of protein liquid-liquid phase separation. Nat. Commun. 12. https://doi.org/10.1038/s41467-021-27433-y.
10. Mohammad-Beigi, H., Mason, T.O., Rovers, T.A.M., Jæger, T.C., Møller, M.S., Ipsen, R., et al. (2026) Taylor dispersion analysis of micellization (TDAM) reveals distinct assembly and dissociation pathways of α-, β-, and κ-casein micelles. Food Hydrocoll. 173. https://doi.org/10.1016/j.foodhyd.2025.112301.
11. Madsen, A.V., Kristensen, P., Buell, A.K. and Goletz, S. (2023) Generation of robust bispecific antibodies through fusion of single-domain antibodies on IgG scaffolds: a comprehensive comparison of formats. MAbs 15.https://doi.org/10.1080/19420862.2023.2189432.
12. Pedersen, M.E., Haegebaert, R.M.S., Østergaard, J. and Jensen, H. (2021) Size-based characterization of adalimumab and TNF-α interactions using flow induced dispersion analysis: assessment of avidity-stabilized multiple bound species. Sci. Rep. 11. https://doi.org/10.1038/s41598-021-84113-z.
13. Pedersen, M.E., Gad, S.I., Østergaard, J. and Jensen, H. (2019) Protein Characterization in 3D: Size, Folding, and Functional Assessment in a Unified Approach. Anal. Chem. 91, 4975-4979. https://doi.org/10.1021/acs.analchem.9b00537.
14. Pinheiro, F., Nowak, J.S., Zueva, E., Pheasant, E.C., Grene, I.K., Lampinen, V., et al. (2025) Screening de novo designed protein binders in unpurified lysate using flow induced dispersion analysis. Protein Science 34. https://doi.org/10.1002/pro.70286.
15. Willmer, P., Hundahl, A.C., Marie, R. and Jensen, H. (2025) Continuous Titration Based Method for Rapid In-Solution Analysis of Non-Covalent Interactions. Chemistry-Methods 5. https://doi.org/10.1002/cmtd.202400059. 16. Norrild, R.K., Mason, T.O., Boyens-Thiele, L., Ray, S., Mortensen, J.B., Fritsch, A.W., et al. (2024) Taylor Dispersion-Induced Phase Separation for the Efficient Characterisation of Protein Condensate Formation. Angew. Chem. Int. Ed.63. https://doi.org/10.1002/anie.202404018.

